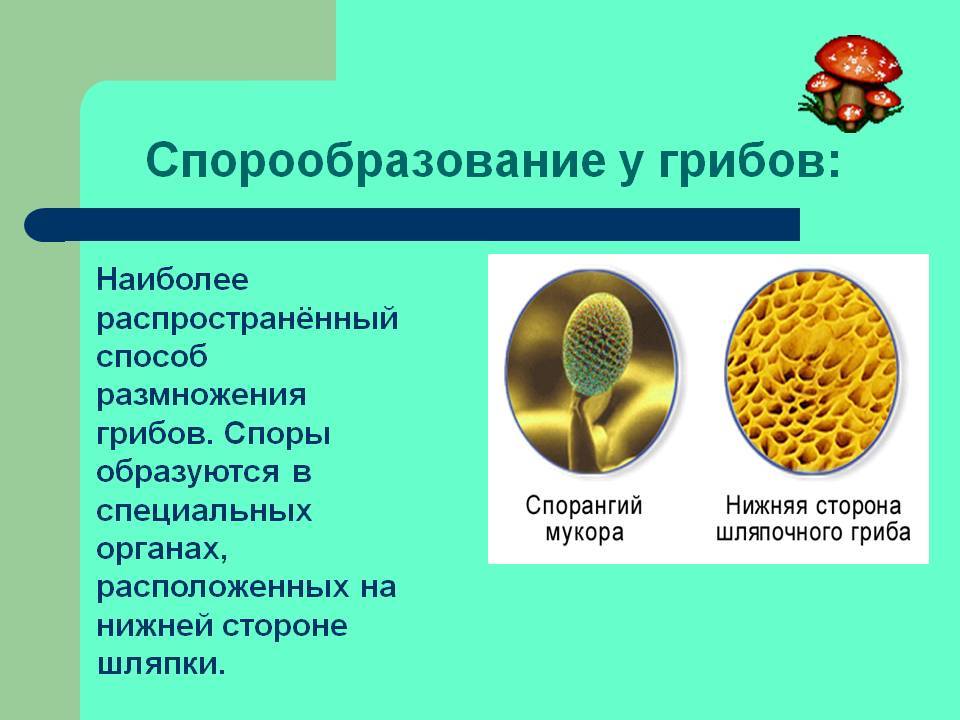
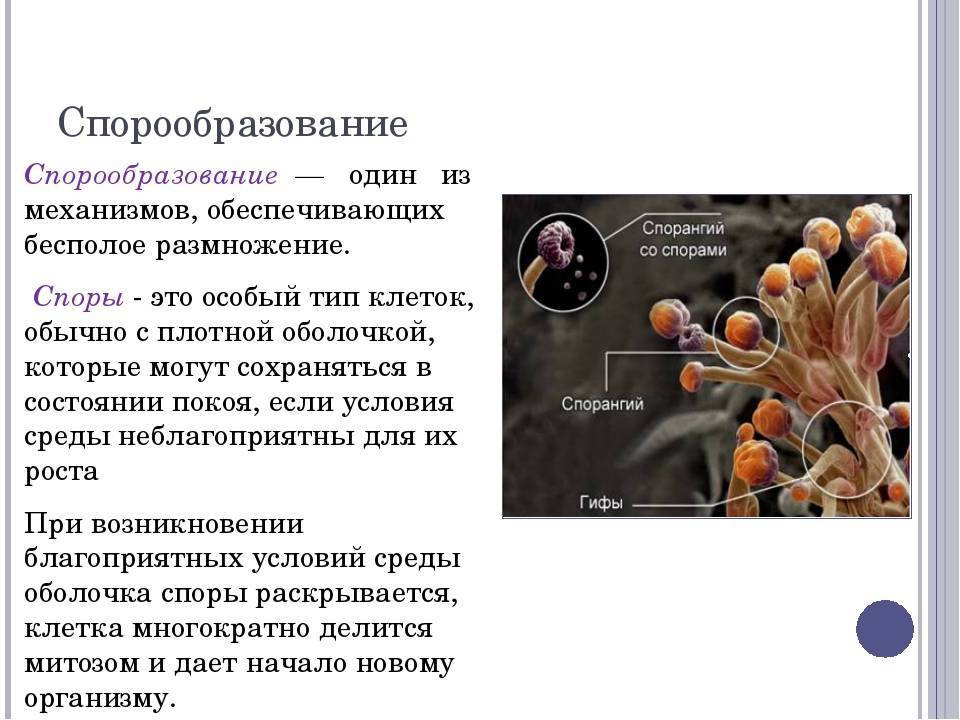
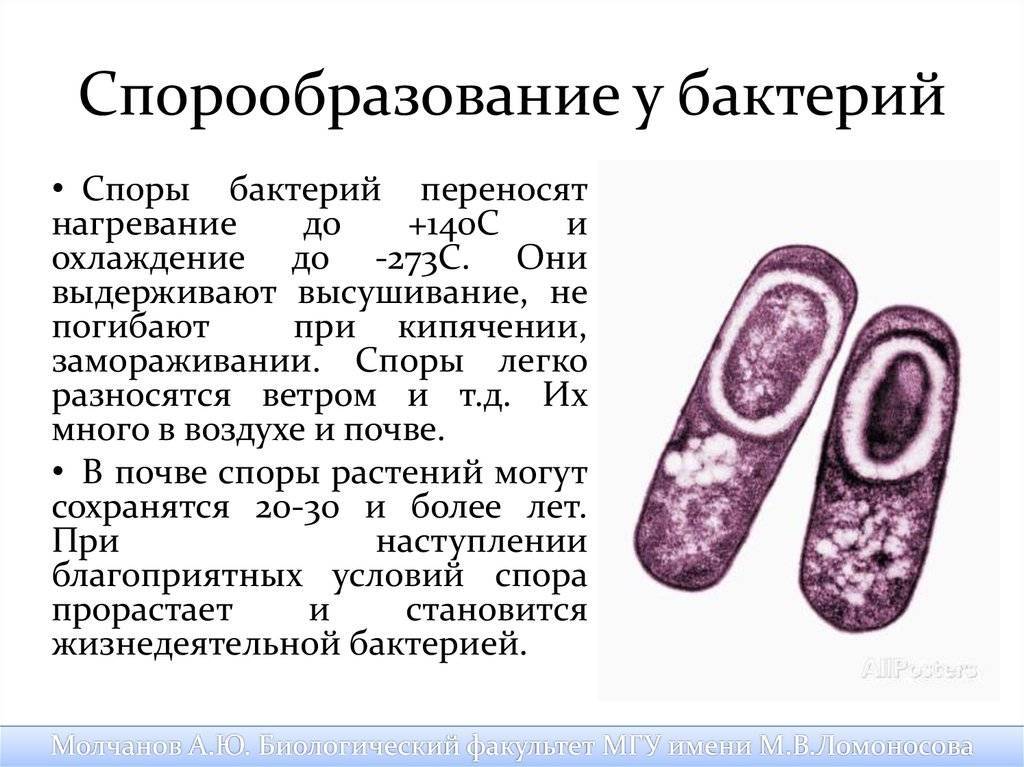
Классификация спор
Споры можно классифицировать несколькими способами:
По спорообразующей структуре
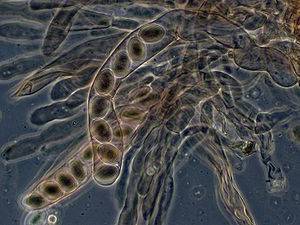
Аски из сморчка высокого , содержащий аскоспоры
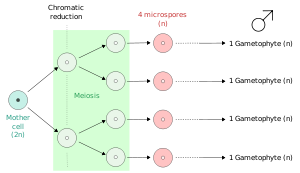
В растениях , микроспоры , а в некоторых случаях мегаспор образуются из всех четырех продуктов мейоза.
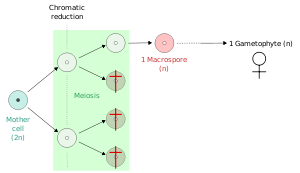
Напротив, у многих семенных растений и гетероспористых папоротников только один продукт мейоза становится мегаспорой (макроспорой), а остальные дегенерируют.
Грибы
У грибов и грибоподобных организмов споры часто классифицируются по структуре, в которой происходит мейоз и образование спор. Так как грибы часто классифицируются в соответствии с их споровым производством структур, эти споры часто характерны для конкретного таксона из грибов .
- Спорангиоспоры : споры, продуцируемые спорангиями многих грибов, таких как зигомицеты .
- Зигоспоры : споры, продуцируемые зигоспорангием , характерные для зигомицетов .
- Аскоспоры : споры, продуцируемые асками , характерные для аскомицетов .
- Базидиоспоры : споры, продуцируемые базидиумом , характерные для базидиомицетов .
- Эциоспоры : споры, продуцируемые эцием некоторых грибов, например ржавчины или головни .
- Урединиоспоры : споры, продуцируемые урединиумом некоторых грибов, таких как ржавчина или головня .
- Телиоспоры : споры, вырабатываемые телиумом некоторых грибов, таких как ржавчина или головня .
- Ооспоры : споры, продуцируемые оогониумом , характерные для оомицетов .
Красные водоросли
- Карпоспоры : споры карпоспорофита , характерные для красных водорослей .
- Тетраспоры : споры, продуцируемые тетраспорофитом , характерные для красных водорослей .
По функциям
- Хламидоспоры : покоящиеся с толстыми стенками споры грибов, созданные для выживания в неблагоприятных условиях.
- Споры паразитических грибов можно разделить на внутренние споры, которые прорастают внутри хозяина, и внешние споры, также называемые спорами окружающей среды, высвобождаемые хозяином для заражения других хозяев.
По происхождению в течение жизненного цикла
-
Мейоспоры : споры, продуцируемые мейозом ; они, таким образом, являются гаплоидными и дают начало гаплоидной дочерней клетке (дочерним клеткам) или гаплоидной особи. Примеры являются клетка — предшественников из гаметофита из семенных растений , найденных в цветах ( покрытосеменные ) или конусов ( голосеменные ), и зооспоры , полученные из мейоза в спорофитах из водорослей , таких как Ulva .
- Микроспоры : мейоспоры, дающие начало мужскому гаметофиту ( пыльца семенных растений).
- Мегаспоры (или макроспоры ): мейоспоры, дающие начало женскому гаметофиту (у семенных растений гаметофит формируется внутри семяпочки ).
- Митоспоры (или конидии , конидиоспоры ): споры, образующиеся в результате митоза ; они характерны для аскомицетов . Грибы, в которых обнаружены только митоспоры, называются «митоспорическими грибами» или «анаморфными грибами» и ранее классифицировались по таксону Deuteromycota (см. Teleomorph, anamorph и holomorph ).
- Зооспоры : подвижные споры, которые перемещаются с помощью одного или нескольких жгутиков , их можно найти в некоторых водорослях и грибах .
- Апланоспоры : неподвижные споры, которые, тем не менее, потенциально могут разрастать жгутики.
- Аутоспоры : неподвижные споры, которые не могут развить жгутики.
- Баллистоспоры : споры, которые принудительно высвобождаются или выбрасываются из плодового тела грибав результате действия внутренней силы, такой как повышение давления. Большинство базидиоспор также являются баллистоспорами, и еще одним ярким примером являются споры рода Pilobolus .
- Статисоспоры : споры, которые выходят из плодового тела грибов в результате воздействия внешней силы, например, капель дождя или проходящего мимо животного. Примеры — клубнички .
Размножение бактерий
Известно много способов размножения, наблюдаемых у различных бактерий.
У подавляющего числа представителей этой группы микроорганизмов размножение осуществляется путем деления клеток на две части.
В средней части физиологически подготовленной к размножению клетки за счет впячивания цитоплазматической мембраны образуется поперечная перегородка.
Расщепляясь, она разделяет клетку на две половинки. Образовавшиеся новые клетки могут быть несколько неодинаковыми по размеру, так как перегородка не всегда проходит посередине материнской клетки.
Кокки в процессе размножения последовательно делятся в одной, двух или трех взаимно перпендикулярных плоскостях.
После деления они остаются в той или иной мере скрепленными друг с другом, в результате чего возникают сочетания кокков, отличающиеся по взаимному расположению (см.рис. 1): диплококки — парные кокки; стрептококки — цепочки кокков; тетракокки — по четыре кокка; сарцины — в форме правильных тючков по 8, 16 шт.; стафилококки — скопления, напоминающие грозди винограда.
При очень слабой связи или ее отсутствии между возникающими при делении клетками образуются микрококки, во взаимном расположении которых нет никаких закономерностей. Они расположены поодиночке или в виде случайных скоплений по несколько экземпляров.
Палочки (бактерии, бациллы), подобно коккам, могут располагаться парами по длине — диплобактерии и цепочками — стрептобактерии.
Большинство же палочек располагается одиночно, беспорядочно. По внешним очертаниям отдельные представители палочковидных заметно отличаются друг от друга. Известны палочки строго цилиндрической формы, бочковидные, с резко обрубленными, вогнутыми или заостренными концами и др.
Размножение делением не сводится только к удвоению числа клеток.
Структурные элементы и вещества материнской клетки еще и перераспределяются между возникающими новыми клетками. Большая часть клеток нового поколения наследует бездефектные структуры родительских организмов, вторая — менее полноценные. В связи с таким распределением по прошествии нескольких циклов деления образуется какое-то количество нежизнеспособных клеток. Установлено, что доля таких клеток, приходящаяся на каждый цикл деления, составляет примерно 10 % общего числа.
Бактерии обладают большой скоростью размножения, которая зависит от условий питания, температуры, доступа воздуха и др.
При благоприятных условиях клетка может делиться через каждые 20—30 мин, т.е. за сутки может произойти 48—72 цикла удвоения.









Споры водорослей
Водоросли производят споры для бесполого размножения. Эти споры могут быть не подвижными (апланоспоры), или подвижными (зооспоры) и перемещаться из одного места в другое с помощью жгутиков. Некоторые водоросли способны размножаться половым или бесполым способом. Когда условия благоприятны, зрелые водоросли делятся и производят споры, которые развиваются в новых особей.
Споры гаплоидные и производятся митозом. Во времена, когда условия неблагоприятны для развития, водоросли подвергаются половому размножению и образовывают гаметы. Эти половые клетки сливаются, чтобы стать диплоидным зигоспорами. Зигоспора останется бездействующей до тех пор, пока условия не станут благоприятными еще раз. В это время зигоспора подвергается мейозу для создания гаплоидных спор.
Некоторые водоросли имеют жизненный цикл, который чередуется между различными периодами бесполого и полового размножения. Этот тип жизненного цикла называется чередованием поколений и состоит из гаплоидной и диплоидной фаз. В гаплоидной фазе структура, называемая гаметофитом, производит мужские и женские гаметы (половые клетки). При слиянии этих гамет образуется зигота. В диплоидной фазе зигота развивается в диплоидную структуру, называемую спорофитом. Спорофит продуцирует гаплоидные споры через мейоз.
Споры растений
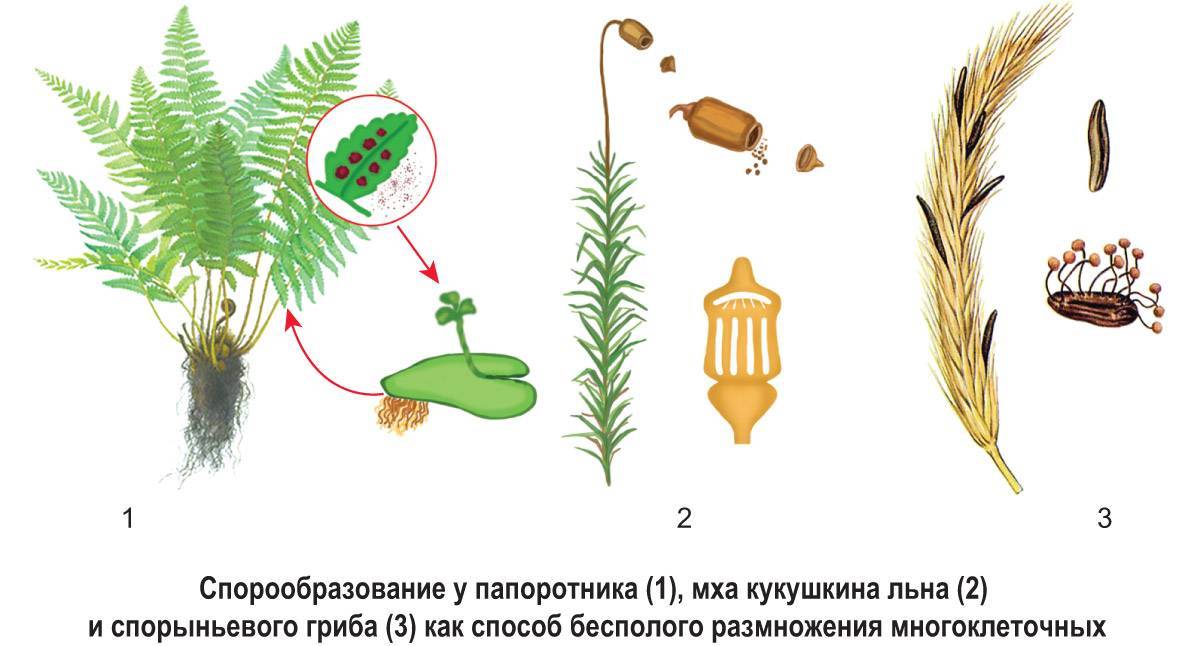
Как водоросли и грибы, растения также демонстрируют чередование поколений. Растения без семян, такие как папоротники и мхи, развиваются из спор. Споры производятся внутри спорангий и выпускаются в окружающую среду. Первичной фазой жизненного цикла несосудистых растений, таких как мхи, является генерация гаметофитов (половая фаза).
Фаза гаметофита состоит из зеленой мшистой растительности, а фаза спорофита (бесполая фаза) состоит из удлиненных стеблей со спорами, заключенными внутри спорангий, расположенных на кончике стеблей. У сосудистых растений, которые не производят семена, такие как папоротники, поколения спорофит и гаметофит являются независимыми. Лист папоротника представляет собой зрелый диплоидный спорофит, тогда как спорангии на нижней стороне листьев образовывают споры, которые развиваются в гаплоидный гаметофит.
У цветковых растений (покрытосеменных) и нецветковых семенных растений генерация гаметофитов полностью зависит от доминантного спорофита. У покрытосеменных, цветок вырабатывает как мужские микроспоры, так и женские мегаспоры. Микроспоры содержатся в пыльце, а мегаспоры производятся внутри цветочной завязи. При опылении микроспоры и мегаспоры объединяются, образуя семена.
Классификация спорообразующих организмов
Растения
Споры сосудистых растений всегда гаплоидны . Сосудистые растения либо homosporous (или isosporous) или разноспоровые . Гомоспористые растения производят споры одного размера и типа.
Гетероспористые растения, такие как семенные растения , колосовидные , иголочки и папоротники отряда Salviniales, производят споры двух разных размеров: более крупная спора (мегаспора) фактически функционирует как «женская» спора, а меньшая (микроспора) функционирует как «споры». мужчина». Такие растения обычно дают начало двум видам спор из отдельных спорангиев: мегаспорангий , продуцирующий мегаспоры, или микроспорангий , продуцирующий микроспоры. У цветковых растений эти спорангии встречаются в плодолистиках и пыльниках соответственно.
Грибы
Споры грибов обычно образуются в результате полового или бесполого размножения. Споры обычно гаплоидны и превращаются в зрелых гаплоидных особей посредством митотического деления клеток ( урединиоспоры и телиоспоры среди ржавчины являются дикариотическими). Дикариотические клетки возникают в результате слияния двух гаплоидных клеток гамет. Среди спорогенных дикариотических клеток кариогамия (слияние двух гаплоидных ядер) происходит с образованием диплоидной клетки. Диплоидные клетки подвергаются мейозу с образованием гаплоидных спор.
Рассредоточение
Споры, выбрасываемые грибами.
У грибов как бесполые, так и половые споры или спорангиоспоры многих видов грибов активно распространяются путем насильственного изгнания из их репродуктивных структур. Этот выброс обеспечивает выход спор из репродуктивных структур, а также перемещение по воздуху на большие расстояния. Таким образом, многие грибы обладают специализированными механическими и физиологическими механизмами, а также структурами на поверхности спор, такими как гидрофобины , для выброса спор. Эти механизмы включают, например, насильственный выброс аскоспор, обусловленный структурой аска, и накопление осмолитов в жидкостях аска, что приводит к взрывному выбросу аскоспор в воздух.
Принудительный выброс отдельных спор, называемых баллистоспорами, включает образование небольшой капли воды ( капли Буллера ), которая при контакте со спорой приводит к высвобождению ее снаряда с начальным ускорением более 10 000 g . Другие грибы полагаются на альтернативные механизмы высвобождения спор, такие как внешние механические силы, примером которых являются клубочки . Привлечение насекомых, таких как мухи, к плодоносящим структурам, благодаря их яркой окраске и гнилостному запаху, для распространения спор грибов — еще одна стратегия, наиболее часто используемая вонючими рогами .
Было показано, что у мха обыкновенного гладкошпончатого ( Atrichum undulatum ) вибрация спорофита является важным механизмом высвобождения спор.
В случае сосудистых растений , распространяющих споры, таких как папоротники , распределение очень легких спор ветром обеспечивает большую способность к их распространению. Кроме того, споры менее подвержены хищничеству животных, чем семена, потому что они почти не содержат запаса пищи; однако они более подвержены грибным и бактериальным хищникам. Их главное преимущество состоит в том, что из всех форм потомства для образования спор требуется меньше всего энергии и материалов.
У шиповника Selaginella lepidophylla распространение частично достигается за счет необычного типа диаспоры — перекати-поле .
Определение
Термин « спора» происходит от древнегреческого слова σπορά spora , означающего « семя , посев», связанного с σπόρος sporos , «сеять», и σπείρειν speirein , «сеять».
Говоря простым языком, разница между «спорой» и « гаметой » заключается в том, что спора прорастает и превращается в спорлинг , в то время как гамета должна объединиться с другой гаметой, чтобы сформировать зиготу, прежде чем развиваться дальше.
Основное различие между спорами и семенами как единицами распространения состоит в том, что споры одноклеточные, первая клетка гаметофита, тогда как семена содержат внутри себя развивающийся зародыш (многоклеточный спорофит следующего поколения), образованный слиянием мужской гаметы пыльцевая трубка с женской гаметой, образованная мегагаметофитом внутри семяпочки. Споры прорастают, давая начало гаплоидным гаметофитам, а семена прорастают, давая начало диплоидным спорофитам.


Внешняя анатомия
Ископаемые споры трилете (синие) и тетрады спор (зеленые) позднесилурийского происхождения
Трехволновая пыльца Ricinus
При большом увеличении споры часто имеют сложные узоры или орнаменты на внешней поверхности. Для описания особенностей таких паттернов была разработана специальная терминология. Некоторые отметки представляют собой отверстия, места, где жесткая внешняя оболочка спор может проникнуть во время прорастания. Споры можно разделить на категории в зависимости от положения и количества этих отметин и отверстий. Споры Alete не показывают линий. У споров монолета на споре имеется единственная узкая линия (laesura). Это указывает на то, что материнская спора разделена на четыре части по вертикальной оси. У спор трилете каждая спора показывает три узкие линии, расходящиеся от центрального полюса. Это показывает, что четыре споры имели общее происхождение и первоначально находились в контакте друг с другом, образуя тетраэдр. Более широкое отверстие в форме канавки можно назвать кольпусом . По количеству кольпи различают основные группы растений. У Eudicots есть трехслойные споры (т. Е. Споры с тремя кольпами).
Споры тетрады и трилете споры
Заключенные в оболочку тетрады спор считаются самым ранним свидетельством растительной жизни на суше, начиная с середины ордовика (ранний Лланвирн, ~ ), периода, из которого еще не были обнаружены макрофоссилии. Отдельные споры трилете, напоминающие споры современных криптогамных растений, впервые появились в летописи окаменелостей в конце ордовикского периода.
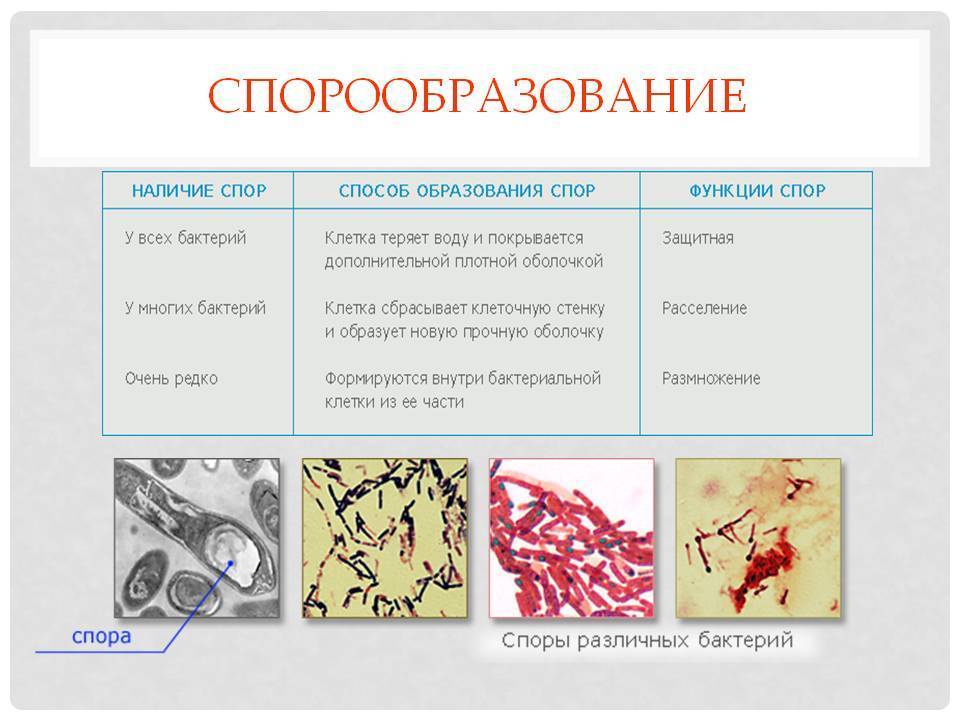
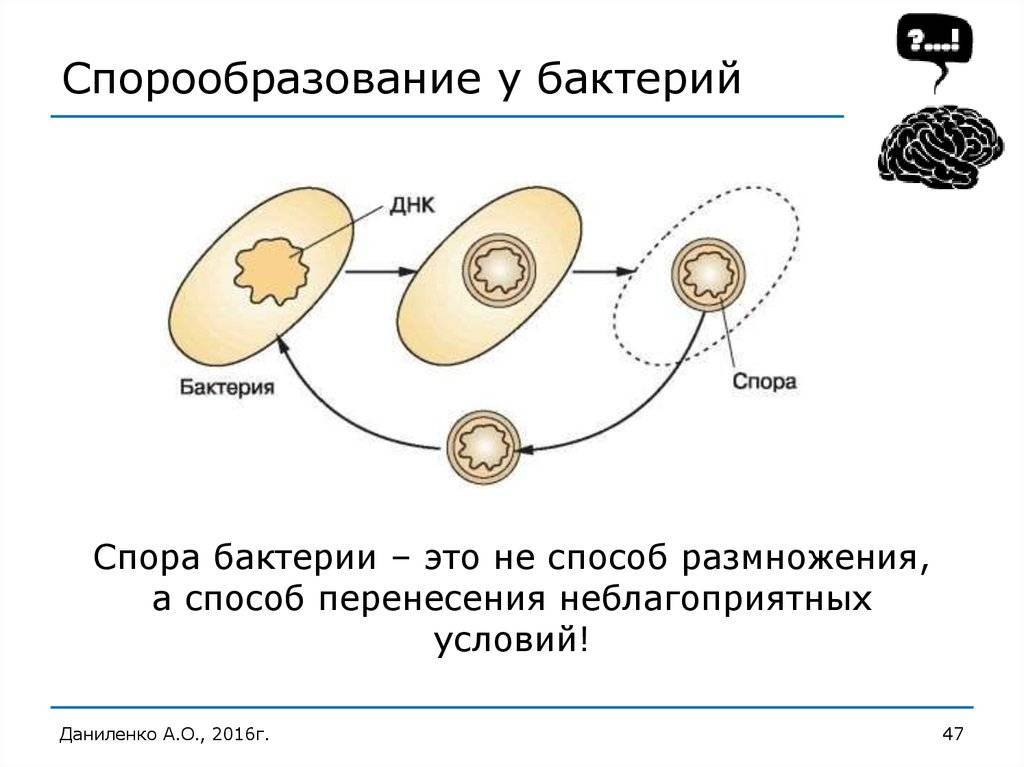
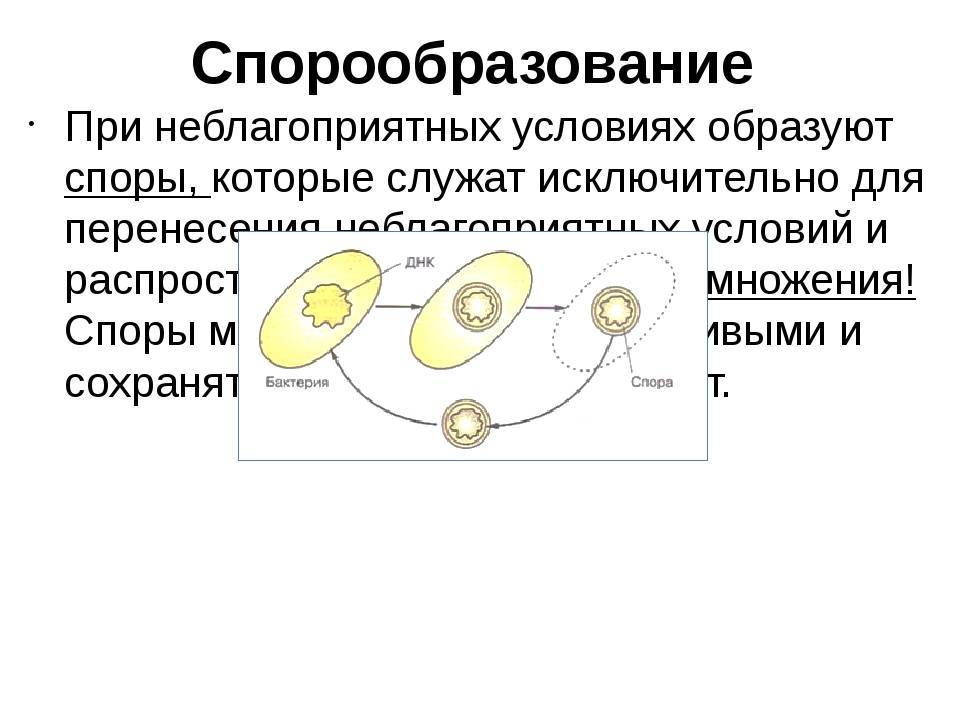
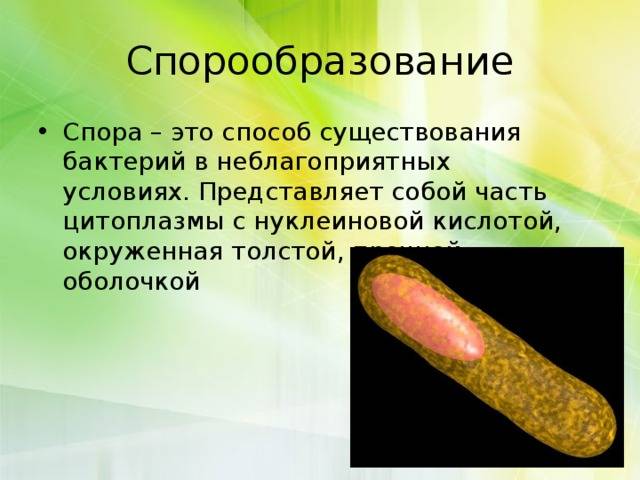
Споры
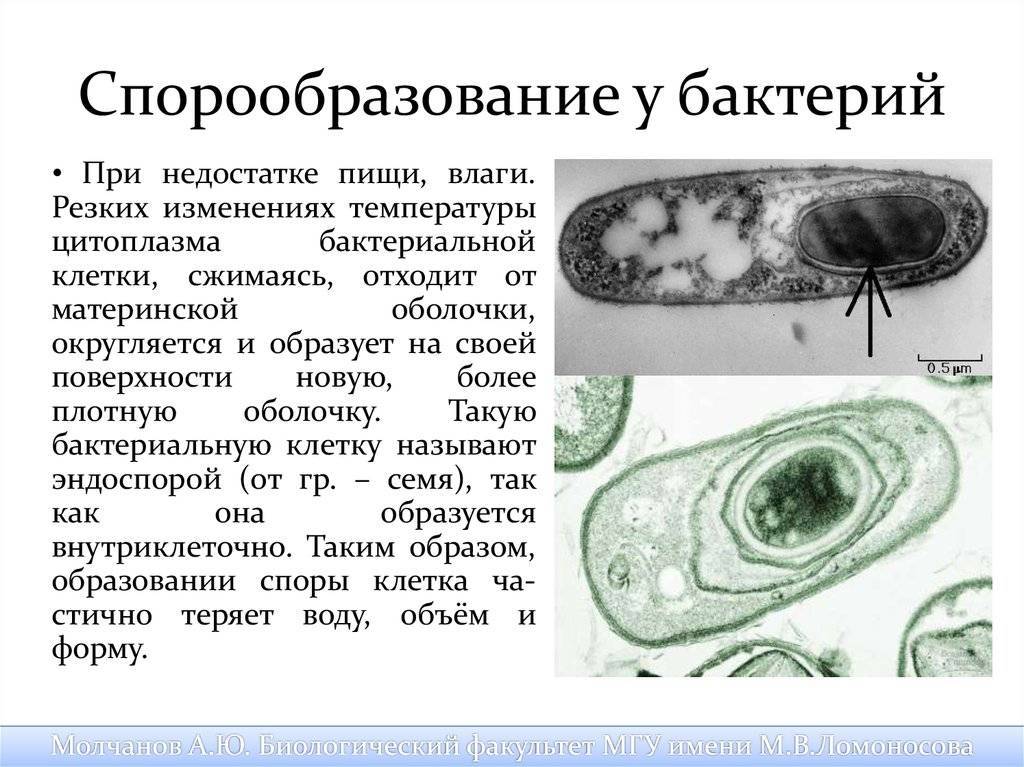
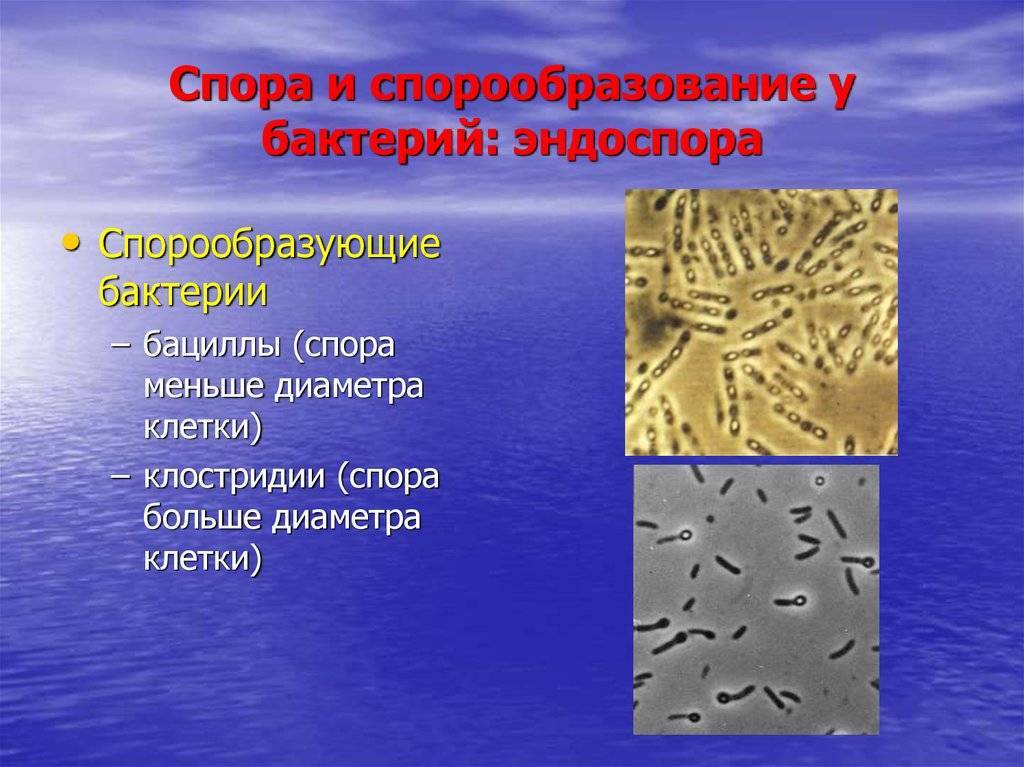
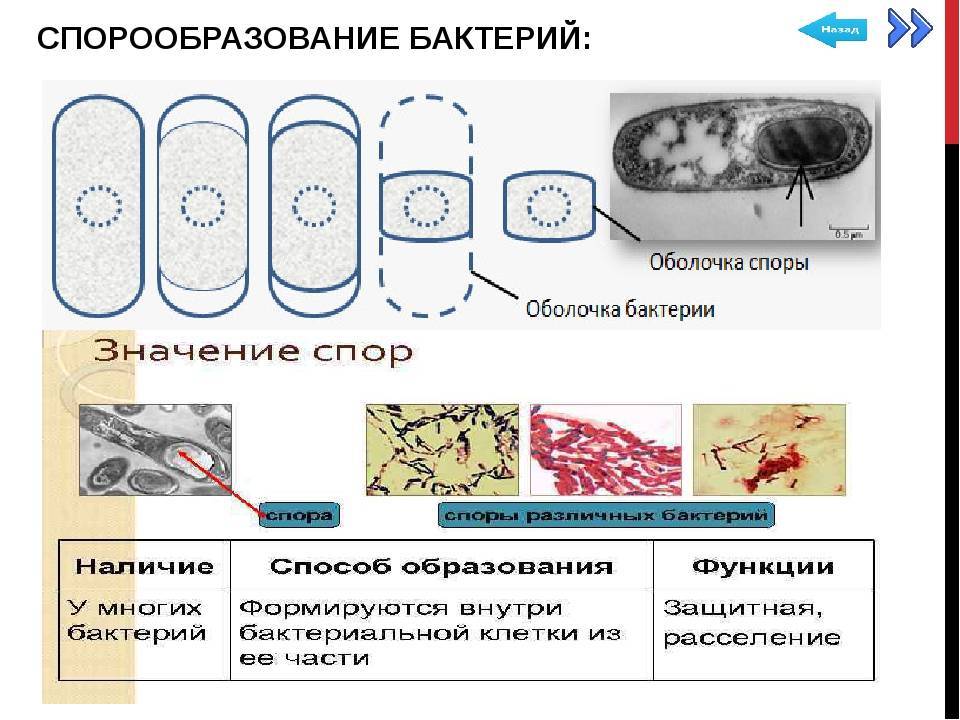
Споры – форма покоящихся грамположительных бактерий. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). При этом внутри одной бактерии образуется одна спора. Образование спор способствует сохранению вида и не является способом размножения. Спорообразующие палочковидные аэробные бактерии, у которых размер споры не превышает диаметр клетки, называются бациллами. Спорообразующие палочковидные анаэробные бактерии, у которых размер споры превышает размер бактериальной клетки, называются клостридиями.
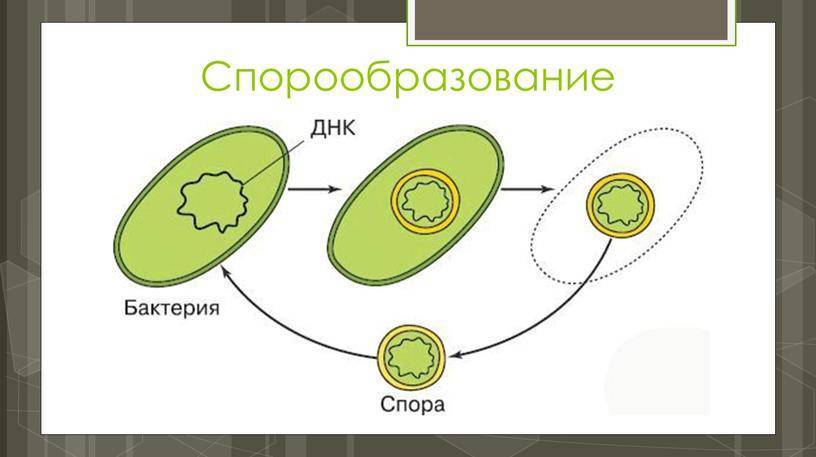
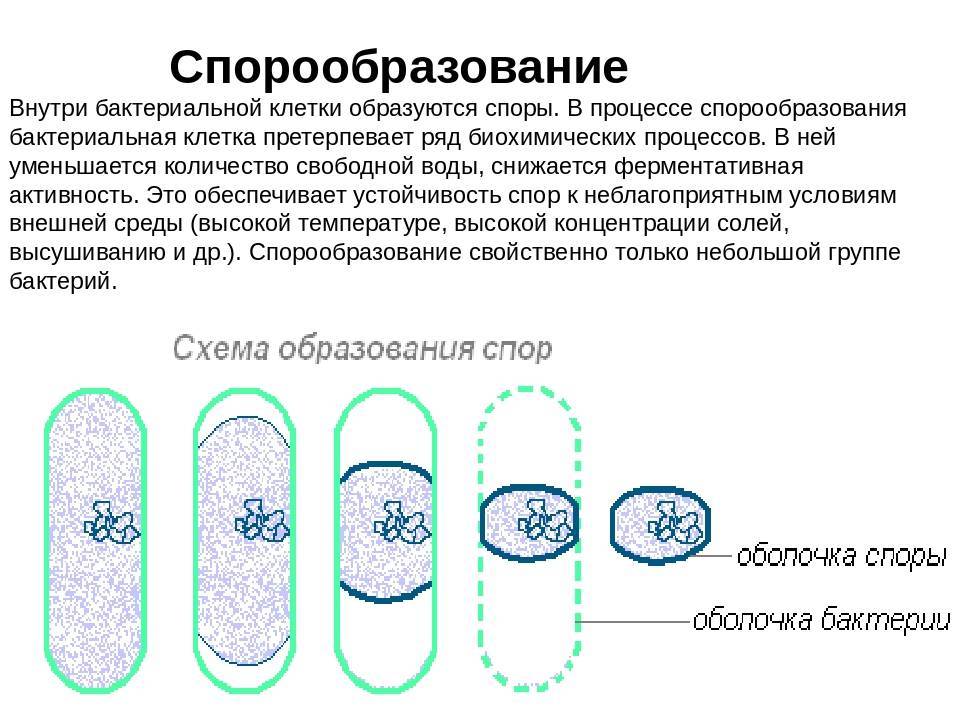
 Схема образования споры (по Г.Шлегелю). А и Б – образование септы. В и Г – окружение протопласта споры мембраной материнской клетки. Д – формирование кортекса и оболочек споры. Е – схема строения зрелой споры: 1 – цитоплазма с нуклеоидом; 2 – ЦМ споры; 3 – клеточная стенка споры; 4 – кортекс; 5 – внутренняя оболочка споры; 6 – наружная оболочка споры; 7 – экзоспориум.
Схема образования споры (по Г.Шлегелю). А и Б – образование септы. В и Г – окружение протопласта споры мембраной материнской клетки. Д – формирование кортекса и оболочек споры. Е – схема строения зрелой споры: 1 – цитоплазма с нуклеоидом; 2 – ЦМ споры; 3 – клеточная стенка споры; 4 – кортекс; 5 – внутренняя оболочка споры; 6 – наружная оболочка споры; 7 – экзоспориум.
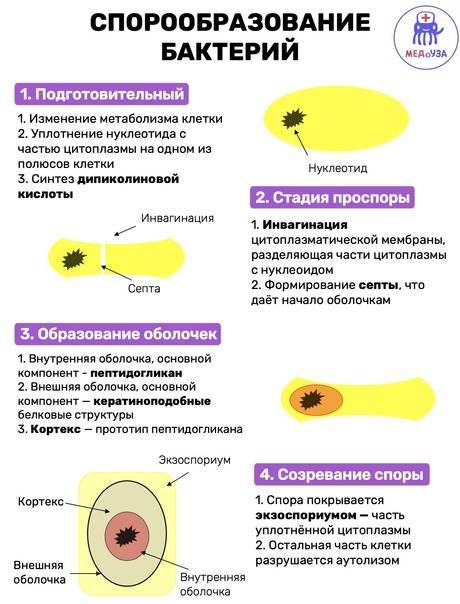
Процесс спорообразования (споруляция) проходит ряд стадий. Вначале на одном из полюсов бактериальной клетки происходит конденсация нуклеоида и отделение его за счет образования септы. Затем ЦПМ начинает обрастать образовавшийся протопласт споры и возникает складка, состоящая из двух слоев ЦПМ, позднее они сливаются, в результате образовавшаяся предспора оказывается окруженной двойной оболочкой. Между обращенными друг к другу мембранами образуется зародышевая стенка, кортекс, а также расположенные снаружи от мембран наружная и внутренняя оболочки.
Протопласт споры содержит ЦПМ, цитоплазму, хромосому, все компоненты белоксинтезирующей системы и анаэробной энергообразующей системы. Стенка споры непосредственно окружает внутреннюю мембрану и представлена пептидогликаном, из которого формируется клеточная стенка прорастающей клетки. Кортекс – самый толстый слой оболочки споры, чувствительный к лизоциму. Оболочка споры построена из кератиноподобного белка. Плохая проницаемость ее определяет высокую устойчивость споры к действию различных химических веществ. Экзоспорий – липопротеиновая оболочка, содержащая немного углеводов. В состав споры входит дипиколиновая кислота, обусловливающая термоустойчивость споры. Затем вегетативная часть клетки отмирает, и спора сохраняется во внешней среде в течение длительных сроков.
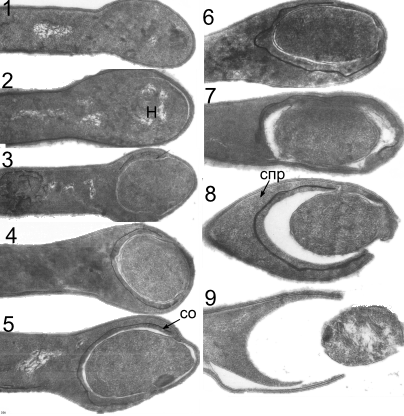
Clostridium tetani. Электронная микроскопия. Стадии образования споры. Н – нуклеоид; СО – споровая оболочка; СПР – спорангий. Х80000.«Авакян А.А., Кац Л.Н., Павлова И.Б. Атлас анатомии бактерий, патогенных для человека и животных. М «Медицина».-1972.-183 с.»
Способность ряда патогенных бактерий образовывать длительно сохраняющиеся во внешней среде споры, обладающие высокой термоустойчивостью, обусловлена низким содержанием воды, повышенной концентрацией кальция, структурой и химическим составом ее оболочки.
В благоприятных условиях споры прорастают, проходя три последовательные стадии: активацию, инициацию, прорастание. При этом из одной споры образуется одна бактерия. Прорастание споры происходит в течение 4-5 ч, в то время как образование споры продолжается 18-20 ч.
Спорообразование, форма и расположение спор в клетке (вегетативной) является видовым свойством бактерии, что позволяет отличать их друг от друга. Форма спор может быть овальной, шаровидной, расположение в клетке – терминальное, т.е. на конце палочки (возбудитель столбняка), субтерминальное – ближе к концу палочки (возбудитель ботулизма) и центральное (сибиреязвенная бацилла).
Споры можно выявить при обычном окрашивании бактериальной клетки в виде не окрашенной области внутри бактериальной клетки и по методу Ожешко.
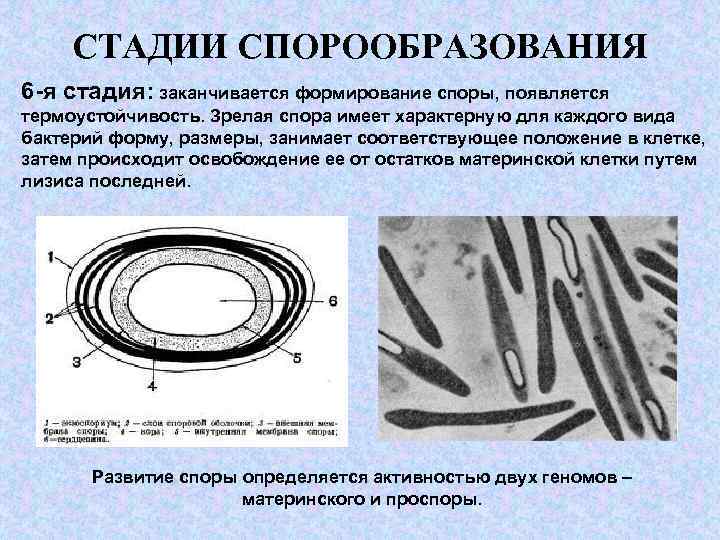
Четвертый этап споруляции – созревание споры
В ходе четвертого этапа бактериальная спора приобретает характерную для вида форму и занимает определенное положение в клетке.
Диаметр бактериальной споры может быть меньше или больше диаметра вегетативной клетки. В результате клетка с созревающей спорой может иметь форму веретена или тенистой ракетки.
Созревающие споры могут располагаться в клетке различным образом:
- центрально – в центарльной части материнской клетки;
- субтерминально – приближенно к одному из концов клетки;
- терминально – в одном из концов бактериальной клетки.
Четвертый этап споруляции заканчивается лизисом материнской клетки, которую в данном случае часто именуют «спорангий».
Третий этап споруляции – образование оболочки
В синтезе оболочки споры принимают участие две мембраны, ограничивающие предспору внутри материнской клетки.
Мембрана протопласта проспоры синтезирует снаружи от себя стенку зародышевой клетки (зародыша).
Мембрана, происходящая от материнской цитоплазматической мембраны, синтезирует вовнутрь кору споры или кортекс, состоящий из многослойного муреина, более кислого, чем муреин клеточной стенки материнской клетки.
Кроме котекса и стенки зародыша, синтезируется наружная оболочка споры, состоящая в значительной степени из полипептидов.
Эндоспоры большинства видов бактерий формируют ещё один слой – экзоспориум. В его состав входят белки, липиды, углеводы.
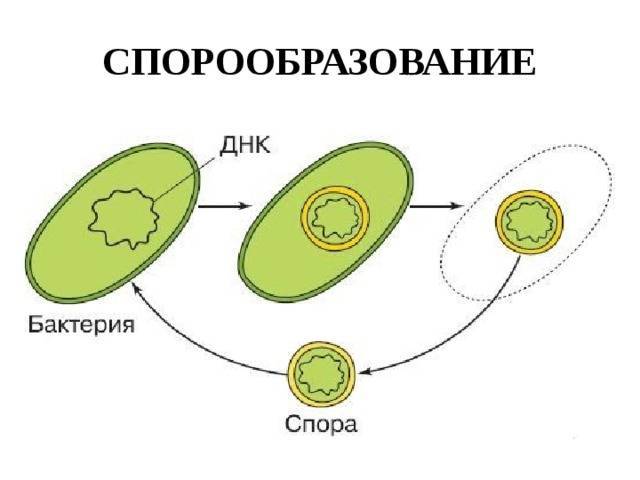




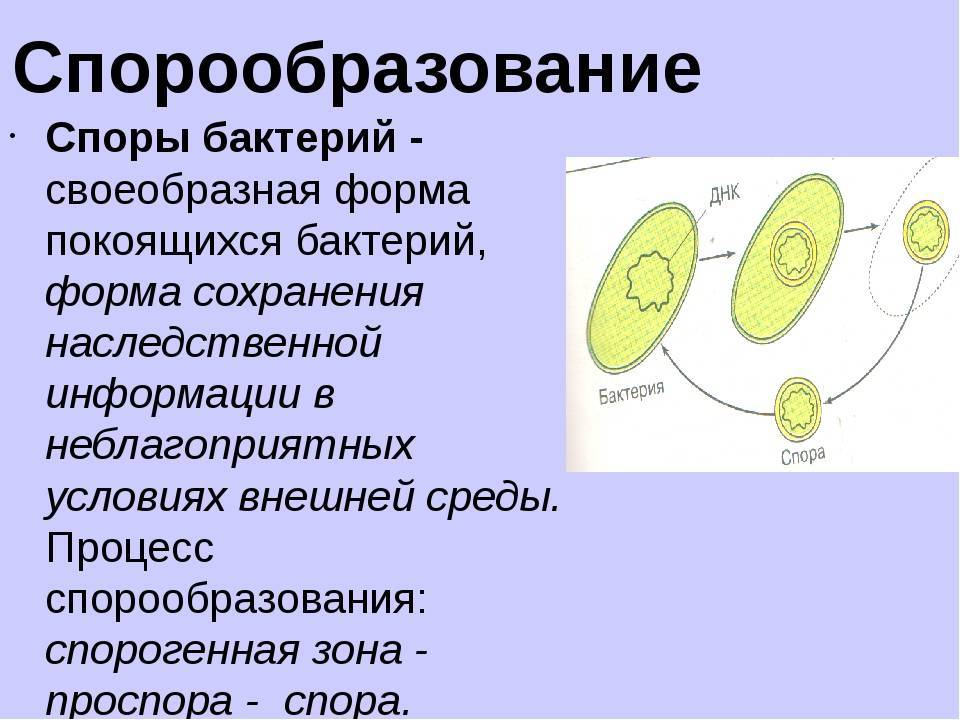
Споры бактерий
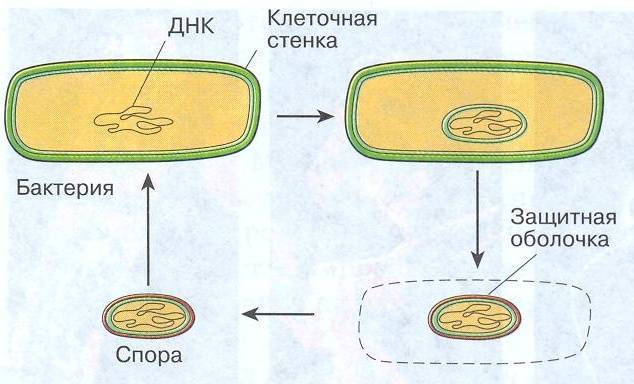
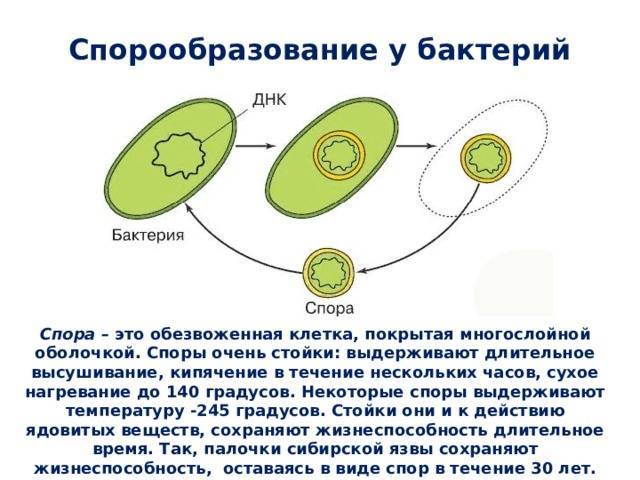
Некоторые бактерии образовывают споры, называемые эндоспорами, как средство борьбы с экстремальными условиями окружающей среде, которые угрожают их выживанию. Эти условия включают высокие температуры, засушливость, наличие токсичных ферментов или химических веществ и отсутствие пищи.
Спорообразующие бактерии развивают толстую клеточную стенку, которая является водонепроницаемой и защищает бактериальную ДНК от высыхания и повреждения. Эндоспоры могут выживать в течение длительных периодов времени, пока условия не изменятся и не станут пригодными для прорастания. Примеры бактерий, способных образовывать эндоспоры, включают клостридии (Clostridium) и бациллы (Bacillus).
Какие микроорганизмы образуют споры
Из одной клетки за это время возникло бы 4714169·1015клеток, через 36 ч микробная масса составила бы около 400 т.
Если бы размножение постоянно проходило с такой скоростью, то из одной клетки в течение 5 дней могло бы образоваться такое количество клеток, что общий объем их оказался бы равным объему всех морей и океанов.
Практически беспрерывного деления микробов не происходит.
Размножению их мешают многие моменты: истощение питательной среды, накопление продуктов собственного обмена и другие физические, химические и биологические факторы внешней среды. Так, при снижении температуры на 10 °С скорость размножения снижается в 2—3 раза.
Попадая в новые условия, на свежий субстрат, микробы не сразу начинают размножаться.
Проходит некоторое время до начала увеличения их числа (фаза задержки роста), в течение которого они приспосабливаются к среде обитания и подготавливают самую среду. После этого начинается бурное размножение, замедляющееся затем по мере исчерпания питательных ресурсов и накопления продуктов жизнедеятельности бактерий в среде.
Быстрое развитие микробиологической порчи продуктов — скисание, окисление, плесневение, гниение и др.
— как раз и объясняется исключительно высокой скоростью размножения бактерий.
Неспорообразующие анаэробы
Какие бактерии образуют споры
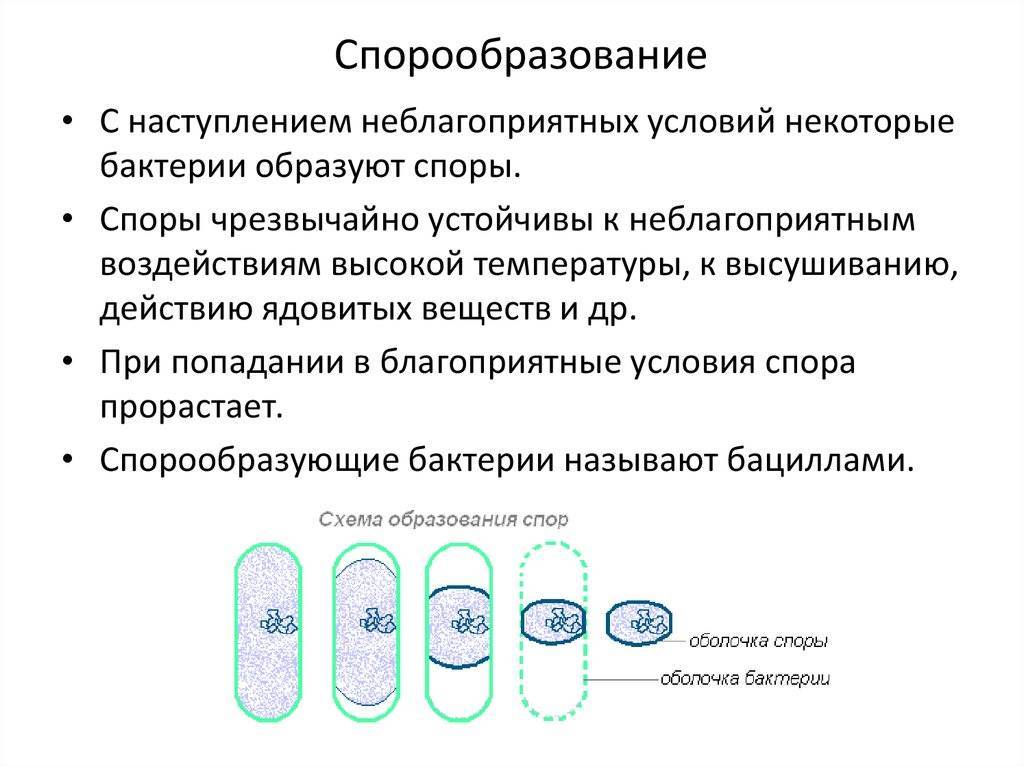
При наступлении для бактерий неблагоприятных условий они способны образовывать споры. Неблагоприятными условиями могут быть отсутствие питательных веществ в среде, изменение ее кислотности, высокие или низкие температуры, пересыхание среды и другое.
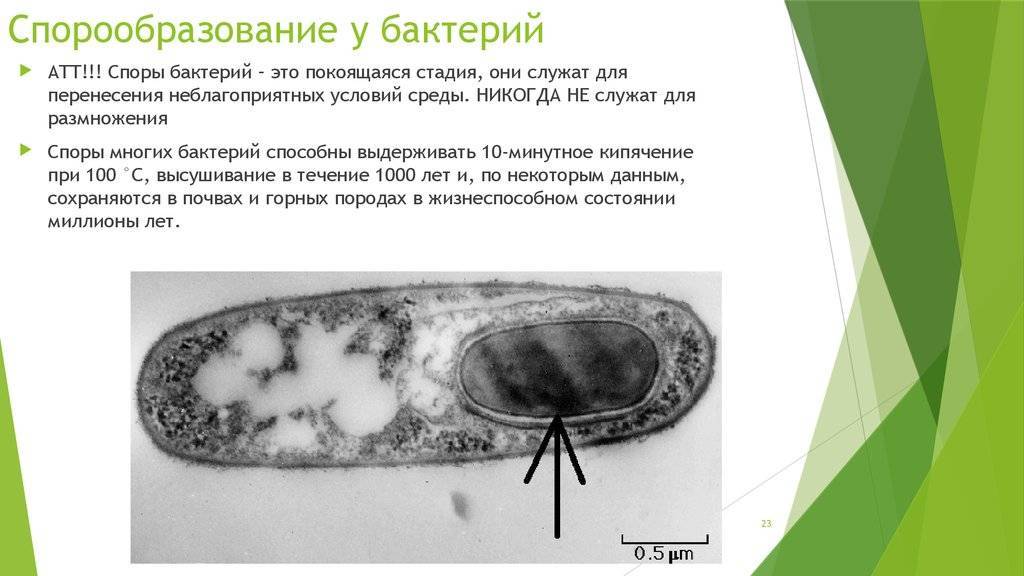
Образование спор бактериями — это в первую очередь способ пережить неблагоприятные условия среды. В отличие от других организмов у бактерий спорообразование почти не используется для размножения.
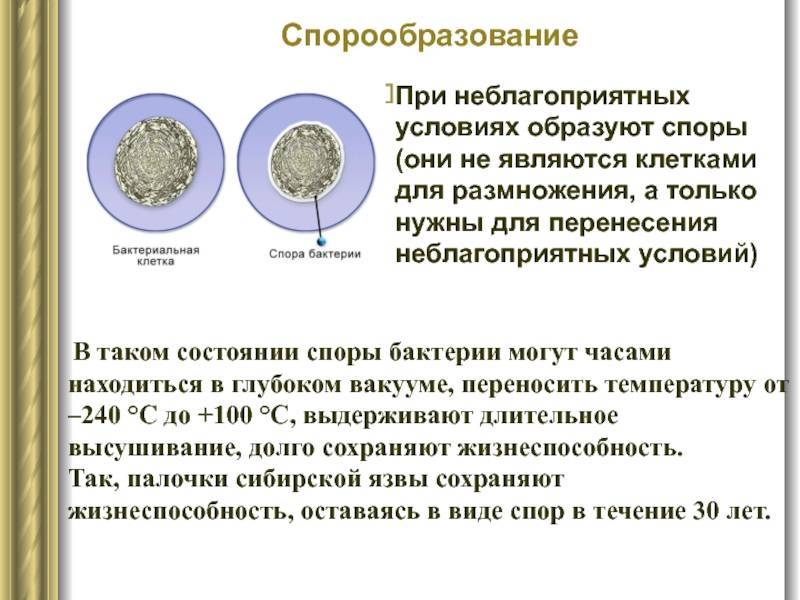
Споры бактерий сохраняют жизнеспособность в весьма неблагоприятных условиях внешней среды. Они способны переживать крайне высокие и низкие температуры, сохранять жизнеспособность на протяжении очень многих лет. Так известны бактерии, споры которых могут прорастать через 1000 лет. У других бактерий споры выдерживают кипячение. Бывает, что споры способны пережить температуру меньше -200 градусов Цельсия.
В те времена, когда жизнь на Земле только появилась, и на ней существовали преимущественно только бактерии, возможно погодные условия могли быстро меняться, становиться весьма суровыми. Чтобы выжить, бактерии эволюционно выработали в себе способность к спорообразованию. На сегодняшний день бактерии могут жить там, где другие организмы выжить не могут.
В спорах бактерий все жизненные процессы почти прекращены, цитоплазмы мало, и она густая. Спора покрыта толстой оболочкой, защищающей ее от разрушающих факторов внешней среды. Однако спора содержит все необходимое (в том числе ДНК бактерии), чтобы в благоприятных условиях прорасти и образовать полноценную бактериальную клетку.

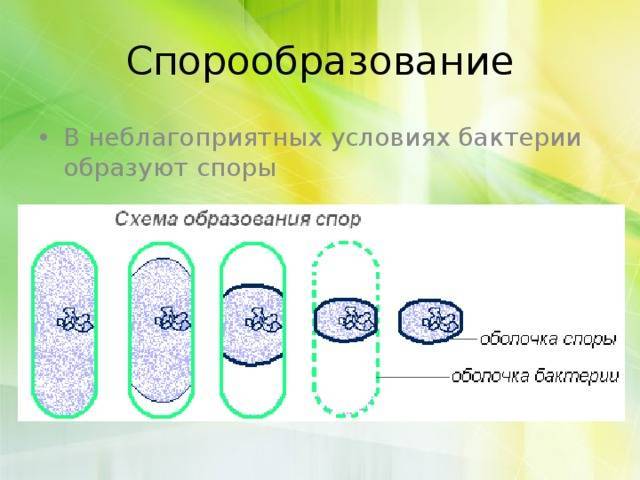
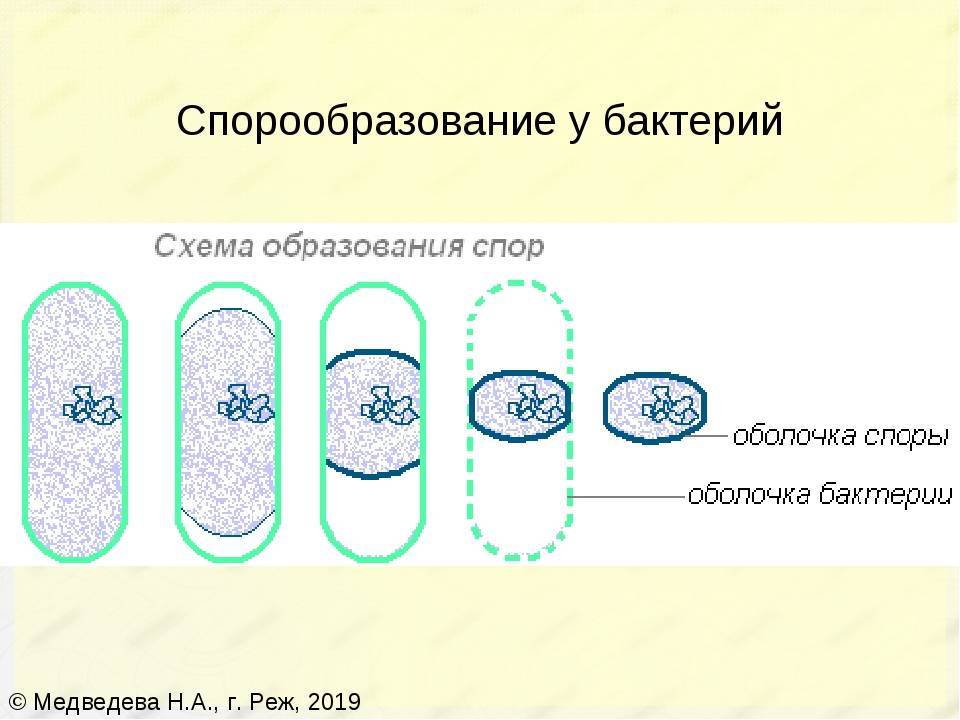
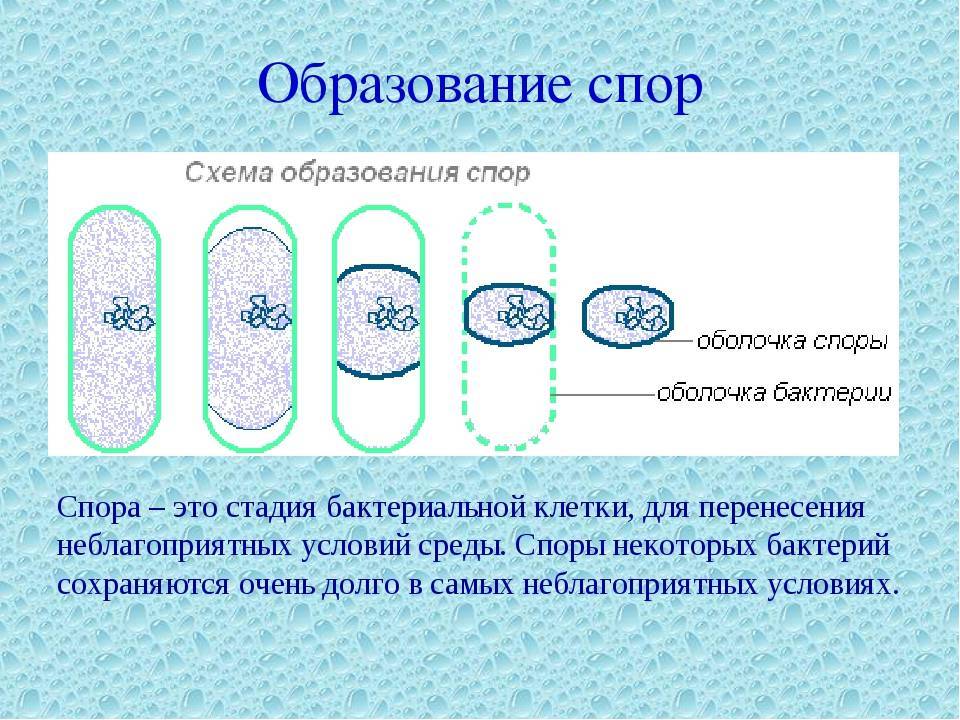
Большинство бактерий образуют споры, которые называют эндоспорами. В основном их образуют палочковидные бактерии. «Эндо» значит «внутри». То есть у большинства бактерий споры образуются внутри клетки. При образовании спор происходит впячивание клеточной мембраны, и внутри бактерии обособляется область — будущая спора. Туда переходит ДНК. Вокруг этой области образуется толстый слой так называемой коры, которая будет защищать спору. С ее внутренней и внешней стороны присутствует мембрана. С внешней стороны от мембраны есть еще несколько оболочек.
У палочковидных бактерий эндоспоры могут образовываться в разных местах клетки. У одних — в середине, у других — ближе к концу, у третьих — у самого края палочки-клетки.
Существуют виды бактерий, которые образуют не эндоспоры, а экзоспоры, цисты и другие формы покоящихся форм. «Экзо» говорит о том, что спора образуется не внутри клетки бактерии, а как бы снаружи от нее. Образование экзоспор происходит путем образования своеобразных почек у клетки. После чего такие почки покрываются толстой оболочкой, превращаются в споры и отделяются.
С помощью спор бактерии не только переживают неблагоприятные условия, но и расселяются, так как споры очень легкие и легко разносятся ветром и водой.
Размножение грибов
Грибы размножаются вегетативным путем и при помощи
спор.
Вегетативное размножение осуществляется кусочками мицелия. Иногда гифы
распадаются на отдельные клетки (оидии), служащие для размножения.
Споры бывают двух видов: бесполые и половые. Образованию бесполых спор
не предшествует слияние клеток. Бесполые споры могут быть внутренними
(эндогенными) и наружными (экзогенными) (рис.19).
Рис.19. Органы бесполого размножения грибов:
а — оидии; б — конидии; в — спорангии; 1 — конидиеносец; 2 — стеригмы;
3 — конидиоспоры; 4 — спорангий; 5 — спорангиоспоры.
Эндогенными спорами являются спорангиоспоры. Они расположены в
спорангиях, сидящих на длинных ножках — спорангиеносцах. Наружными
спорами являются конидиоспоры. Они расположены цепочками на ножках —
конидиеносцах. Конидиеносцы имеют разную форму. Верхушка может быть
булавовидно утолщена (Aspergillus — аспергиллус), разветвлена
(Реnicillium — пенициллиум, Botrytis — ботритис); конидиоспора может
превратиться в зооспорангий, в котором образуются многочисленные
зооспоры (споры, снабженные жгутиками).
Половые споры образуются в результате слияния двух клеток. Слияние
может быть изогамное (слияние одинаковых клеток) и оогамное (слияние
женской и мужской половых клеток). В первом случае возникает зигоспора,
или зигота, во втором — ооспора.
У грибов наблюдается своеобразный половой процесс, состоящий в слиянии
ядер (кариогамия). Процесс начинается с двухъядерной клетки (дикарион).
После слияния ядер начинается деление слившегося ядра на 4 или 8 спор
(рис.20). Таким путем образуются аскоспоры и базидиоспоры. Аскоспоры
являются внутренними спорами. Они образуются по восемь в сумках или
асках. Базидиоспоры являются наружными спорами и возникают по четыре,
каждая на ножке (стеригмы).
Рис.20. Органы полового размножения грибов:
а — зигота; б — сумка (аск); в — базидия.
У большинства грибов имеются и бесполые, и половые споры, и только
грибы, относящиеся к классу несовершенных грибов, лишены половых спор.
У грибов, имеющих оба вида спор, наблюдается чередование бесполого и
полового способа размножения. Бесполые споры менее устойчивы к
неблагоприятным факторам, поэтому они образуются при благоприятных
условиях и служат для распространения вида. Половые споры более
устойчивы. Они образуются в меньшем количестве и служат для сохранения
вида (например, для перезимовывания).
У некоторых грибов для размножения служат склероции – твердые
образования из уплотненных гиф — и хламидоспоры –
уплотненные,
покрытые толстой оболочкой участки гиф. Эти образования достаточно
устойчивы и способны перезимовывать.